
- APPUNTI PER UNA RICOSTRUZIONE DELLA PREISTORIA LIGURE
- ORIGINI GEOLOGICHE E PRIMI ABITANTI LIGURI
- * * *
- 150 MA
- IL TRIASSICO IN ITALIA: UN MARE TROPICALE
- Estratto da Pregliasco Michele, www.digilands.it.
- * * *
- ORIGINE GEOLOGICA della LIGURIA
- Estratto da Barbara Coppo, www.vegiazena.it.
- * * *
- GEOARCHEOLOGIA DELL’AREA URBANA GENOVESE
- Estratto da Maria Cristina Bonci, Marco Firpo, Caterina Ottomano, in Genova dalle origini all’anno mille, a cura di Melli P., SAGEP, 2014.
- * * *
- HOMO ERECTUS (IN LIGURIA)
- Estratto da Guido Zunino, www.vegiazena.it.
- * * *
- HOMO SAPIENS NEANDERTHAL (IN LIGURIA)
- Estratto da Guido Zunino, www.vegiazena.it.
- * * *
- L’HOMO SAPIENS NEANDERTHAL AI BALZI ROSSI
- GROTTE DI FINALE LIGURE
- GROTTE di TOIRANO
- * * *
- FAUNA NEL PALEOLITICO
- * * *
APPUNTI PER UNA RICOSTRUZIONE DELLA PREISTORIA LIGURE
ORIGINI GEOLOGICHE E PRIMI ABITANTI LIGURI
* * *
150 MA
IL TRIASSICO IN ITALIA: UN MARE TROPICALE
Estratto da Pregliasco Michele, www.digilands.it.
Il mondo 250 milioni di anni fa [MA] era molto diverso: tutti i continenti erano riuniti in una unica grande massa continentale, la pangea, attorno al quale si estendeva il mare di pantalassa.
E’ questo il periodi di tempo che i geologi chiamano il Triassico e nel quale l’Italia non c’era, o meglio, quella porzione di crosta terrestre che sarebbe diventata la penisola italiana, si trovava sommersa da un mare chiamato Tetide ad una latitudine molto vicina all’equatore.
Se con la macchina del tempo ci trasferissimo nell’Italia di quell’epoca troveremmo un mare tropicale costellato da bassi fondali , dalle acque calde e pulite, dal quale emergevano isole e scogliere coralline, mentre in lontananza vedremmo le terre emerse.
Sulle scogliere, a poca profondità, prosperavano spugne, alghe calcaree e coralli, organismi con caratteristiche molto importanti:
- Molti sono organismi costruttori, cioè proteggono il loro corpo molle con un esoscheletro fatto di carbonato di calcio.
- Vivono in simbiosi con un’ alga fotosensibile per cui tendono a vivere a poca profondità, a non più di 30-40 metri, per sfruttare il più possibile la luce del sole .
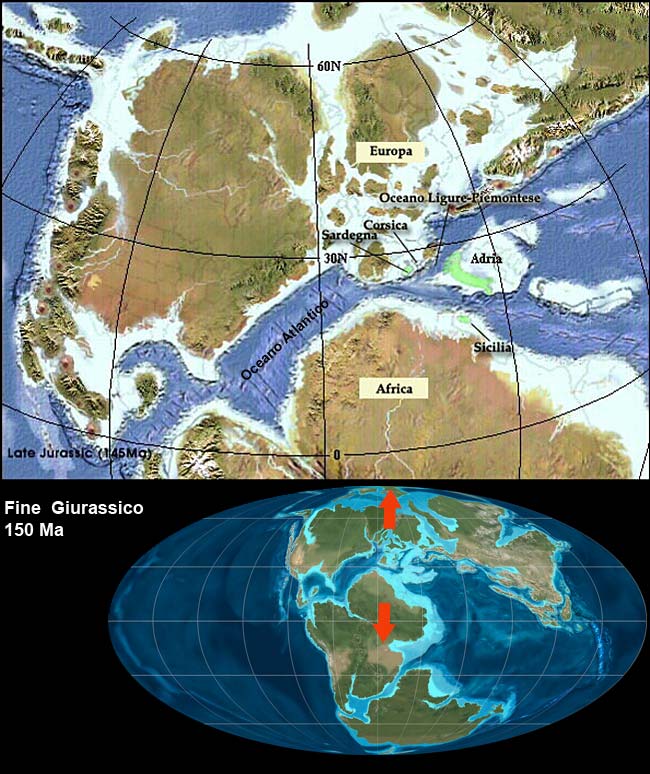
Ron Blakey, NAU Geology (http://cpgeosystems.com/paleomaps.htlm) (modificato da M.Pregliasco) tardo giurassico (150 MA)
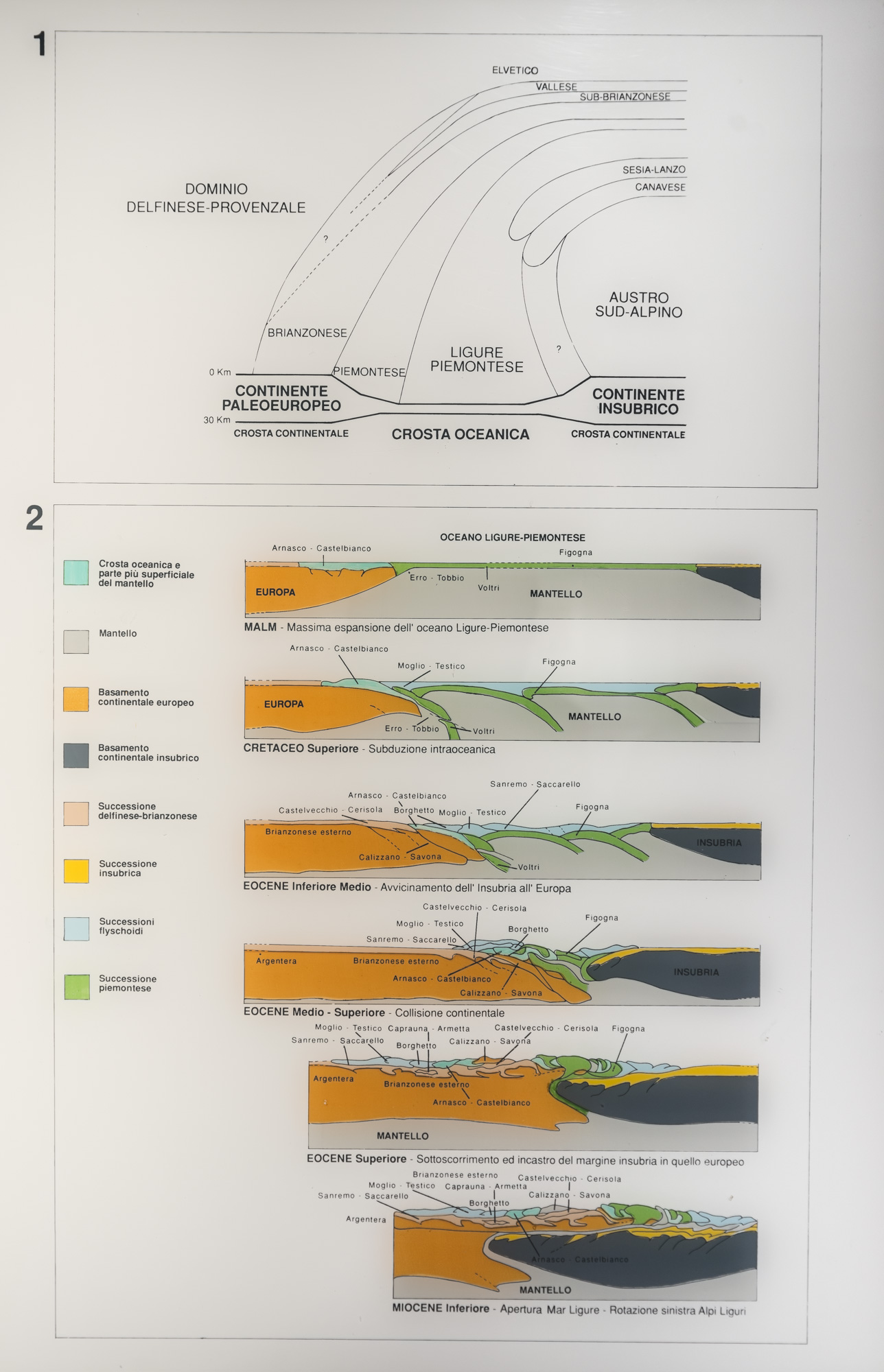
Le fenditure che alla fine del Triassico interessano il centro di Pangea si allargarono sempre di più fino a che, nel Giurassico Medio 180-160 MA, l’America si separa dall’Africa: nasce l‘Oceano Atlantico Centrale.
Questo fenomeno ha una importante conseguenza sull’ area mediterranea: Africa ed Europa cominciano a separarsi e in mezzo nasce l’Oceano Ligure-Piemontese. Nascono così due nuovi margini continentali separati dal neonato Oceano Ligure-Piemontese: l’Adria ad oriente e la costa Europea ad occidente.
L’Adria contiene la maggior parte dei futuri territori italiani (anche se sono sotto i livello del mare) mentre, Sicilia e Sardegna, sono dislocati sul margine africano il primo e su quello europeo il secondo, come evidenziato in verde sulla cartina. Sempre ad Adria appartengono il Mare Adriatico, e parte della ex Iugoslavia e della Grecia, le alpi orientali (comprese: Innsbruck, Salisburgo, Vienna).
* * *
ORIGINE GEOLOGICA della LIGURIA
Estratto da Barbara Coppo, www.vegiazena.it.
Si può affermare che il territorio ligure è stato generato da un meccanismo tettonico affascinante che spiega anche la peculiare varietà dei suoi ambienti. A tutti è nota l’origine recente, geologicamente parlando, dell’Italia, ma forse non molti sanno che circa 225 milioni di anni fa, nel periodo triassico dell’era secondaria, al posto della Liguria si trovava un grande mare, l’Oceano Ligure, e le coste che si fronteggiavano al di qua e al di là dell’acqua erano le terre francesi del Delfinato e quelle toscane che per la loro origine più antica e la loro posizione sono considerate formazioni autoctone.
La Liguria si è originata dalla compressione di questo fondo oceanico per opera di forze tettoniche, le stesse che sono responsabili dell’ apertura dell’oceano Atlantico.
Proverò di seguito a spiegarne il processo analizzando gli eventi in ordine cronologico
e aiutandomi con disegni schematici.
210 milioni di anni fa. Triassico superiore. Tra le terre francesi meridionali e quelle
della pianura toscana si è formata una spaccatura in corrispondenza della quale si ha sprofondamento del suolo e contemporanea risalita di materiale magmatico: è una fossa tettonica simile a quella che oggi possiamo osservare nella Rift-Valley Africana.
Questa depressione è naturalmente coperta da acqua marina (fig. 1).
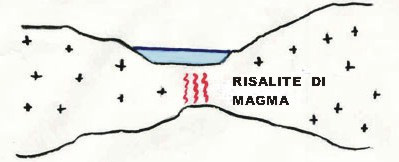
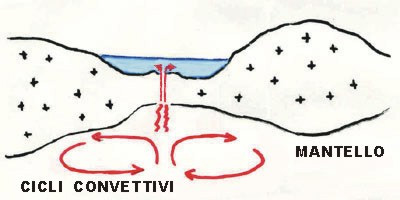
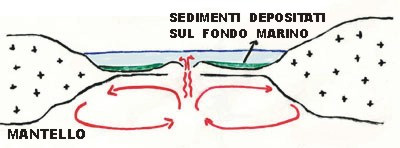
Fig. 2a e 2b.
65 milioni di anni fa. Cretaceo inferiore. L’apertura di una nuova dorsale oceanica, quella atlantica, ad ovest di quella ligure, determina la diminuzione dell’emissione di magma al centro dell’oceano ligure: il suo fondale arresta l’espansione. (fig.3)
 Fig. 3.
Fig. 3.
CRETACEO SUPERIORE: La forte entità della nuova dorsale atlantica determina l’entrata in subduzione del fondale oceanico ligure: in corrispondenza del contatto tra zolla autoctona continentale francese e fondale marino si crea una profonda spaccatura
e il fondale oceanico costituito da materiale a maggiore densità scivola sotto al continente francese, scende fino alle profondità del mantello e fonde. (figg. 4a e 4b)
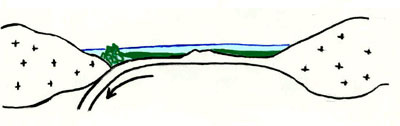

Fig. 4a e 4b.
PALEOCENE E EOCENE: Tutto il materiale sedimentato in milioni di anni sul fondo dell’oceano ligure, essendo più leggero del substrato basaltico che lo sostiene, non sprofonda nella fossa di subduzione ma si accumula sopra ripiegandosi e metamorfosandosi variamente. (fig.5)
 Fig. 5.
Fig. 5.
Si possono vedere nella pratica gli effetti di questa compressione, analizzando le formazioni rocciose della regione. A partire da Voltri e verso ovest si osserva una formazione di rocce scure, dai colori olivastri: sono le serpentiniti, formate dal fondo basaltico che non è riuscito a scendere nel mantello.
Andando verso est si incontrano rocce variamente ripiegate e leggermente metamorfiche
formate da materiale di varia origine sedimentato sull’antico fondo oceanico: sono i calcari ad elmintoidi, sui quali è costruita buona parte della città di Genova, le arenarie del monte Ramaceto (a nord-est di Portofino), i conglomerati del monte di Portofino.
Le vestigia della vecchia dorsale oceanica ligure si possono rinvenire nella falesia di Framura dove sono ben visibili delle formazioni tondeggianti dette Pillows, espressione dell’attività magmatica sottomarina. In definitiva, la Liguria è uno straordinario concentrato di formazioni oceaniche compresse poi una sull’altra dalle forze tettoniche.
Non c’è quindi da stupirsi se nelle formazioni sedimentarie si rinvengano tracce fossili, in alcuni casi così numerosi da dare il nome alla roccia stessa.
————————————
Nel Pliocene inferiore (da 5 a 3 milioni di anni fa) un mare tropicale, la cui linea di riva
si situava all’interno della regione, ricoprì il basamento calcareo. Con il passare del tempo
si accumularono sul fondo di questo mare sedimenti fini e gusci calcarei e silicei di microrganismi marini nelle zone più distanti dalla costa, mentre nelle aree meno profonde, il cui fondo era caratterizzato da sabbie e vaste praterie di posidonia oceanica, vivevano squali, dugonghi (mammiferi erbivori sim ili alle foche) e molluschi con conchiglie anche di grandi dimensioni. (Melli P., Genova preromana, F.lli Frilli ed., 2008)
———————————–
 L’Italia intorno a due milioni di anni fa (a sinistra), quando gran parte
L’Italia intorno a due milioni di anni fa (a sinistra), quando gran parte
della penisola doveva ancora emergere, e durante il picco della
glaziazione del Würm (a destra), circa 18.000 anni fa, quando
l’abbassamento del livello marino fino a circa 120 metri espose territori
oggi sommersi. Tra le due immagini intervengono il sollevamento
tettonico, le variazioni del clima, i terremoti, l’attività dei vulcani.
(Gianluca Valensise, www.museocapellini.it)
——————————–
 Cartello al Museo dei Balzi Rossi (Ventimiglia). Su concessione del Ministero dei Beni e delle Attività Culturali e del Turismo, Sopraintendenza Archeologica della Liguria.
Cartello al Museo dei Balzi Rossi (Ventimiglia). Su concessione del Ministero dei Beni e delle Attività Culturali e del Turismo, Sopraintendenza Archeologica della Liguria.
* * *
GEOARCHEOLOGIA DELL’AREA URBANA GENOVESE
Estratto da Maria Cristina Bonci, Marco Firpo, Caterina Ottomano, in Genova dalle origini all’anno mille, a cura di Melli P., SAGEP, 2014.
La città storica occupa un anfiteatro naturale, intorno all’insenatura in cui si è sviluppata la parte più antica dell’arca portuale, delimitato dagli spartiacque di Granarolo a nordovest e del Righi a nordest, che lo separano dai bacini dei torrenti Polcevera a ponente e Bisagno a levante: si tratta di un’area grossomodo a triangolo isoscele il cui vertice si colloca in corrispondenza del Forte Sperone. Questo anfiteatro è modellato dalle incisioni di undici rivi, oggi tutti coperti, che vanno a sfociare in una baia di forma asimmetrica chiusa alle estremità dai due promontori della Lanterna e del Molo. La costa orientale è più movimentata, con brevi tratti quasi pianeggianti digradanti fino al mare e si prolunga nella piccola penisola del Molo. La parte occidentale, ossia il promontorio della Lanterna spianato nella seconda metà dell’ottocento, è invece costituita da uno sperone montuoso con morfologie via via più acclivi fino a formare una ripida scogliera sul mare. Aldilà dei due spartiacque si allargano le piane alluvionali del Bisagno e del Polcevera.
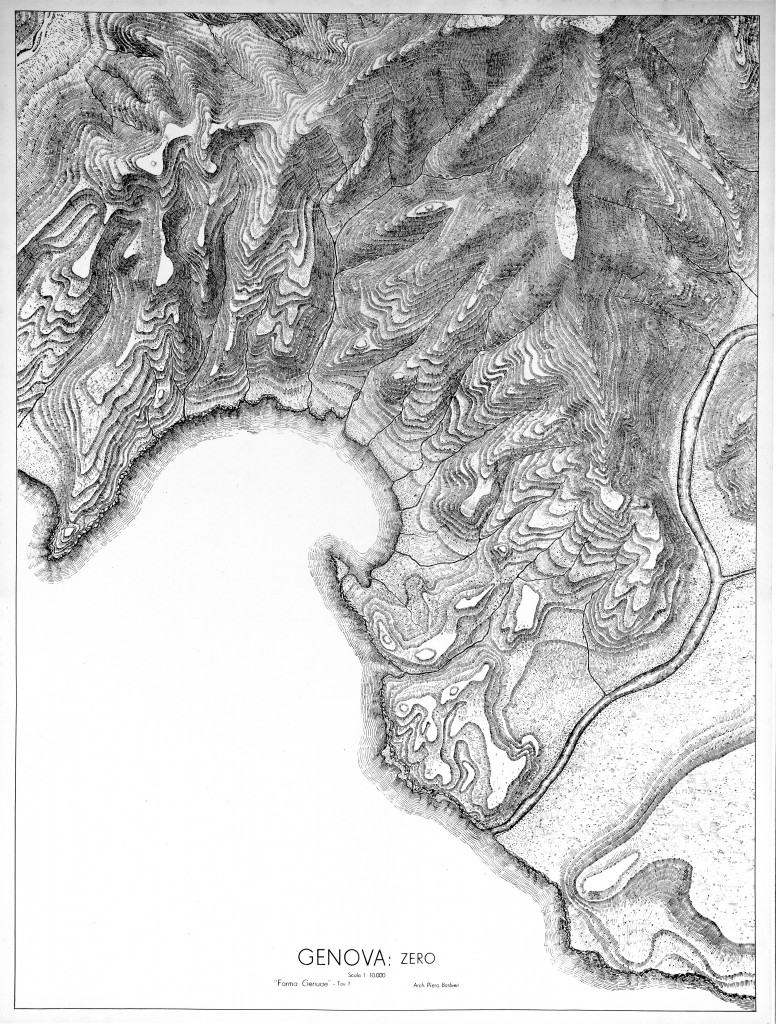 (Barbieri P., Forma Genuae, Municipio di Genova, 1938)
(Barbieri P., Forma Genuae, Municipio di Genova, 1938)
Questa caratteristica morfologia è strettamente connessa con la geologia e la tettonica dell’area. Il substrato geologico dell’anfiteatro di Genova è costituito da rocce appartenenti a tre diverse formazioni: le argilliti di Montoggio, la formazione del Monte Antola e le argille di Ortovero [valle Arroscia, Albenga].
Le argilliti di Montoggio, datate al Campaniano (un intervallo di tempo del Cretaceo Superiore circa tra 84 e 70 milioni di anni fa) e rappresentate da argilliti nere e verdastre depositatesi in mare profondo, sono il litotipo meno diffuso, dato che costituiscono nell’area un unico modesto affioramento nella zona di Granarolo.
La formazione del Monte Antola, la cui genesi risale al Campaniano Superiore (circa tra 77 e 70 milioni di anni fa), è rappresentata da strati alternati di calcareniti, marne e marne calcaree alternate ad argilliti di mare profondo in strati centimetrici; stratigraficamente è successiva alle argilliti di Montoggio sopra citate e costituisce la maggior parte dell’area. Questa potente successione (si ipotizza uno spessore di circa 1000 m) è fortemente deformata. I sedimenti che hanno originato queste rocce si sono depositati ed accumulati in ambiente di piana abissale. Le rocce appartenenti a tale formazione hanno contribuito massicciamente all’edificazione della città, soprattutto durante il Medioevo, attraverso l’utilizzo della nota Pietra di Promontorio, una varietà molto scura e compatta che veniva cavata appunto nell’area del Promontorio della Lanterna [San Benigno]: basta pensare, per esempio, ai portici di Caricamento e alle liste nere del classico parato genovese bianco e nero di molte chiese e palazzi.
Le argille di Ortovero, datate al Pliocene inferiore (circa tra 5 e 4 milioni di anni fa), sono rappresentate da argille marnose, marne, siltiti e arenarie fini di colore grigio azzurro e ricoprono la formazione del Monte Antola in un’area allungata compresa tra piazza Principe, il Porto Antico (zona Calata Gadda), il centro cittadino (piazza De Ferrari, via XX Settembre) fino a San Martino, via Isonzo.
È interessante notare che, dato il contrasto di permeabilità tra i calcari marnosi cretacei (permeabili perché fratturati) e le marne plioceniche (impermeabili o poco permeabili), le zone di contatto tra le due formazioni possono essere caratterizzate dalla presenza di emergenze idriche.
Negli scavi effettuati tra fine ‘800 e inizi ’900 per il riordino urbanistico del centro di Genova sono stati trovati numerosi resti fossili, in ottimo stato di conservazione, di organismi marini, che hanno consentito di ricostruire l’ambiente di genesi di queste rocce, rivelando sotto l’abitato cittadino le testimonianze di un antico mare tropicale. All’inizio del Pliocene circa 5 milioni di anni fa l’area genovese viene ricoperta dal mare, che avanza sulla terra emersa penetrando più all’interno rispetto alla situazione attuale. Sul fondo di questo mare, in zone moderatamente profonde (tra 600 e 1200 m di profondità) si depositavano i sedimenti fini all’origine delle argille di Ortovero e i resti delle strutture scheletriche sia di microorganismi calcarei e silicei sia di molluschi e altri invertebrati di fondi fangosi. A questi si mescolavano i resti di organismi trasportati dalle zone di piattaforma interna, tipici quindi di acque più basse, caratterizzate a volte da fondi sabbiosi o anche da praterie algali o fanerogame marine. Troviamo, infatti, una ricca associazione a molluschi, spesso con conchiglie massicce e di grandi dimensioni, tipici sia di acque basse che di acque più profonde, a cui si aggiungono denti di squali e resti scheletrici di un Sirenide, Metaxytherium subappenninum, un grande mammifero marino erbivoro simile agli attuali Dugonghi. Tutto il complesso faunistico e floristico indica un mare di tipo tropicale-subtropicale.
 Il mare tropicale pliocenico. Ricostruzione grafica di un settore della piattaforma marina continentale di fronte a Genova nel Pleistocene (5-3,5 milioni di anni fa) (Disegno di Fabrizio Boccardo)
Il mare tropicale pliocenico. Ricostruzione grafica di un settore della piattaforma marina continentale di fronte a Genova nel Pleistocene (5-3,5 milioni di anni fa) (Disegno di Fabrizio Boccardo)
—————————————

 “Metaxytherium subappenninum″ di Luca Oddone – Museo Geopaleontologico GAMPS. Con licenza CC BY-SA 3.0 tramite Wikimedia Commons – https://commons.wikimedia.org/wiki/File:Metaxytherium_subappenninum4.JPG#/media/File:Metaxytherium_subappenninum4.JPG
“Metaxytherium subappenninum″ di Luca Oddone – Museo Geopaleontologico GAMPS. Con licenza CC BY-SA 3.0 tramite Wikimedia Commons – https://commons.wikimedia.org/wiki/File:Metaxytherium_subappenninum4.JPG#/media/File:Metaxytherium_subappenninum4.JPG
Metaxytherium subapenninum, Bruno 1839, rappresenta l’unica specie di sirenio che viveva nel mar Mediterraneo nel Pliocene; scomparve definitivamente dal Mediterraneo intorno a 3 milioni di anni fa per il progressivo raffreddamento climatico. Il genere Metaxytherium compare all’inizio del Miocene e mostra un’ampia radiazione geografica: si ritrova fossile nel Mediterraneo, nel Nord e Sud Atlantico e nelle coste orientali del Pacifico. Resti fossili di Metaxytherium subapenninum sono stati rinvenuti, a partire dalla seconda metà dell’Ottocento, in Piemonte, Emilia-Romagna, Liguria e Toscana. Denominato anche vacca marina, il dugongo è un animale di grandi dimensioni che può raggiungere i tre metri di lunghezza, appartiene all’ordine dei Sirenia insieme al lamantino. Sono animali esclusivamente acquatici a dieta erbivora che vivono in acqua marina (dugongo) o anche dolce (lamantino). Il nome dell’ordine, Sirenia, deriva dalla lontana somiglianza che questi esseri hanno con le creature mitologiche per metà donne e per metà pesci. Effettivamente l’espressione del muso e le mammelle posizionate in zona toracica riportano lontanamente alla figura umana. La parte caudale ha poi forma di pinna di pesce per cui la fantasia degli antichi ha sicuramente trovato terreno fertile a questo proposito, anche se i parametri di bellezza con cui si può definire una sirena mitologica non sono certo sovrapponibili a quelli utilizzabili per la descrizione di questo particolare animale. Le dimensioni del dugongo sono notevoli sia per quanto riguarda la lunghezza che il peso che può aggirarsi intorno alla tonnellata . (https://it.wikipedia.org/wiki/Metaxytherium_subapenninum)
* * *
HOMO ERECTUS (IN LIGURIA)
Estratto da Guido Zunino, www.vegiazena.it.
L’ evoluzione umana inizia alcuni milioni di anni fa ed è difficile, in base ai ritrovamenti conosciuti attualmente, datare l’esatto passaggio dai pre-ominidi (scimmie) agli antenati comuni (AUSTRALOPITHECUS) e poi al genere propriamente umano (HOMO).
La teoria evolutiva attualmente più accreditata che spiega questa transizione si riferisce a grandi eventi geologici a livello planetario che hanno interessato l’emisfero Boreale a partire dal MIOCENE (17.000.000 a.C) e sino al PLIOCENE (2.300.000 a.C.).
Per la deriva dei continenti la placca africana venne spinta contro la placca asiatica e contemporaneamente allo spostamento dell’INDIA verso nord, che urtando l’ ASIA formerà la catena dell’ HIMALAYA, in Africa orientale, dall’urto con la penisola arabica, si formerà l’ ALTOPIANO ETIOPICO.
A seguito di questa nuova situazione il flusso delle correnti umide occidentali che portavano la pioggia in Africa orientale viene rallentato. Contemporaneamente il clima cambia diventando più freddo e riducendo ulteriormente le piogge.
L’accumulo di acqua nei ghiacciai (durante le molte glaciazioni di questo periodo) porterà ad una maggior aridità di ampie zone e sommandosi con una minor quantità di pioggia produrrà una parziale desertificazione portando in alcune parti dell’Africa un cambiamento della vegetazione del territorio.
Ad est dell’altopiano Etiopico inizia quindi a cambiare la vegetazione che da arborea diventa arbustiva trasformando le locali foreste d’alto fusto in savane con pochi alberi distanti tra loro, molta erba e bassi arbusti.
Questa modifica ambientale costringe, per pura sopravvivenza, alcune delle razze di scimmie arboricole ivi esistenti a non poter più vivere sugli alberi (che iniziano a mancare) e ad adattarsi alla vita nella savana e nella prateria dove trovano solo rari alberi o bassi arbusti ed erba.
L’altitudine dell’ ALTOPIANO ETIOPICO (da 2.000 ad oltre 4.000 metri s.m.), la depressione della GREAT RIFT VALLEY (una profonda spaccatura che da GIBUTI arriva fino al MALAWI), le pareti scoscese della FOSSA GALLA, il MAR ROSSO, il MAR ARABICO e l’ OCEANO INDIANO dall’altro lato renderanno molto difficile il passaggio delle scimmie da una zona all’altra di quel territorio, diversificandole così nell’evoluzione e costringendo quelle che vivevano nella zona della savana-prateria a vivere a terra, a diventare bipedi in modo permanente per meglio utilizzare gli arti superiori, a diventare carnivori e con una alimentazione più ricca di proteine ad avere più energie e a sviluppare più intuito .
Il bipedismo porta ad un maggior controllo del territorio perchè raddrizzandosi si estende il campo visivo (in un terreno ora quasi sgombro da alberi) e poi permette con le “mani” di raccogliere frutti e bacche selvatici o di trasportare le prede catturate ai punti di incontro con altri suoi simili coabitanti il territorio.
Non veloci nei movimenti, privi di corazze protettive, senza artigli o zanne, l’unico modo per sopravvivere (difendersi ed alimentarsi) sarà quello di impugnare con gli arti superiori, come utensili d’aiuto, rami secchi o pietre trovate casualmente sul terreno.
Molto tempo dopo il passaggio successivo sarà quello di modificare questi utensili rudimentali migliorandoli , ossia strofinandoli sulle rocce ruvide per appuntirli o batterli per scheggiarli e renderli più taglienti.
Nascerà così l’ Homo abilis.
L‘ Homo erectus si posiziona quindi nella scala dell’evoluzione umana dopo l’ Homo abilis (da 2.500.000 a 1.800.000 anni fa) e prima dell’Homo sapiens (200.000/35.000 anni fa).
———————————-
950.000-900.000 AF
Grotta di Vallonet (Francia, tra Roquebrune e Cap Martin). La grotta di Vallonet è al momento il più antico abitato in grotta attualmente conosciuto in Europa. Lo studio delle faune rinvenute nei sedimenti archeologici (in particolare resti di elefanti, ippopotami, bovidi, cervidi, suidi) ha permesso di attribuire al giacimento un’età compresa fra 1,3 e 0,7 milioni di anni mentre lo studio del paleomagnetismo del riempimento della grotta la colloca all’episodio di Jaramillo, tra 0,95 e 0,9 milioni di anni. L’industria litica comprende strumenti su ciottolo e su scheggia. (Museo di Mentone) (www.antiqui.it, www.hominides.com)
Sempre in Francia, l’associazione faunistica e la microfauna della Grotta del Vallonnet (Nizza) permettono un inquadramento di questo complesso, da cui provengono alcuni manufatti su ciottoli di calcare e qualche scheggia, verso la fine del Pleistocene inferiore (episodio paleomagnetico di Jaramillo). Al momento attuale, la Grotta del Vallonnet rappresenta una delle testimonianze meno discutibili di presenza umana in Europa intorno a 1-0,9 MA. (www.treccani.it)
——————————-
Riguardanti un breve periodo intermedio (300.000 – 200.000 anni fa) si troveranno anche le tracce dell’ Homo erectus pre-sapiens, che rappresenta l’anello di congiunzione tra le due genealogie.
I primi ritrovamenti che testimoniano la comparsa dell’ Homo erectus sono databili a circa 1.800.000 anni fa nell’Africa orientale, da dove si diffonderà successivamente sia in Asia che in Europa con lente migrazioni fino a 90.000 anni fa.
In Europa la sua diffusione sarà limitata solo alla fascia centro-meridionale corrispondente agli attuali stati di: Spagna, Francia, Germania, Italia, Grecia e Turchia, e databile tra 1.700.000 e 1.200.000 anni fa. In Italia (più antico sito europeo) resti dell’Homo erectus sono stati rinvenuti a Pirro nord sopra Foggia con datazione attribuibile tra 1.300.000 e 1.700.000 anni fa. Il più antico cranio fossile dell’ H.E. trovato in Italia è stato rinvenuto a Ceprano (Frosinone), mentre ad Isernia (Molise) resti ossei sono stati datati attorno ai 700.000 anni fa. Ed in Spagna, ad Atapuerca, resti umani dell’ H.E. sono stati attribuiti a 1.200.000 anni fa.
La limitazione della diffusione dell’ Homo Erectus solo in queste zone sud-europee era dovuta al fatto che questi erano gli unici territori lasciati liberi dalla spessa coltre dei ghiacci che in quel periodo ricoprivano tutto il nord Europa e metà dell’Asia (glaciazioni di: Donau, Günz, Minde, Riss e Würm da 3.000.000 a circa 10.000 anni fa).
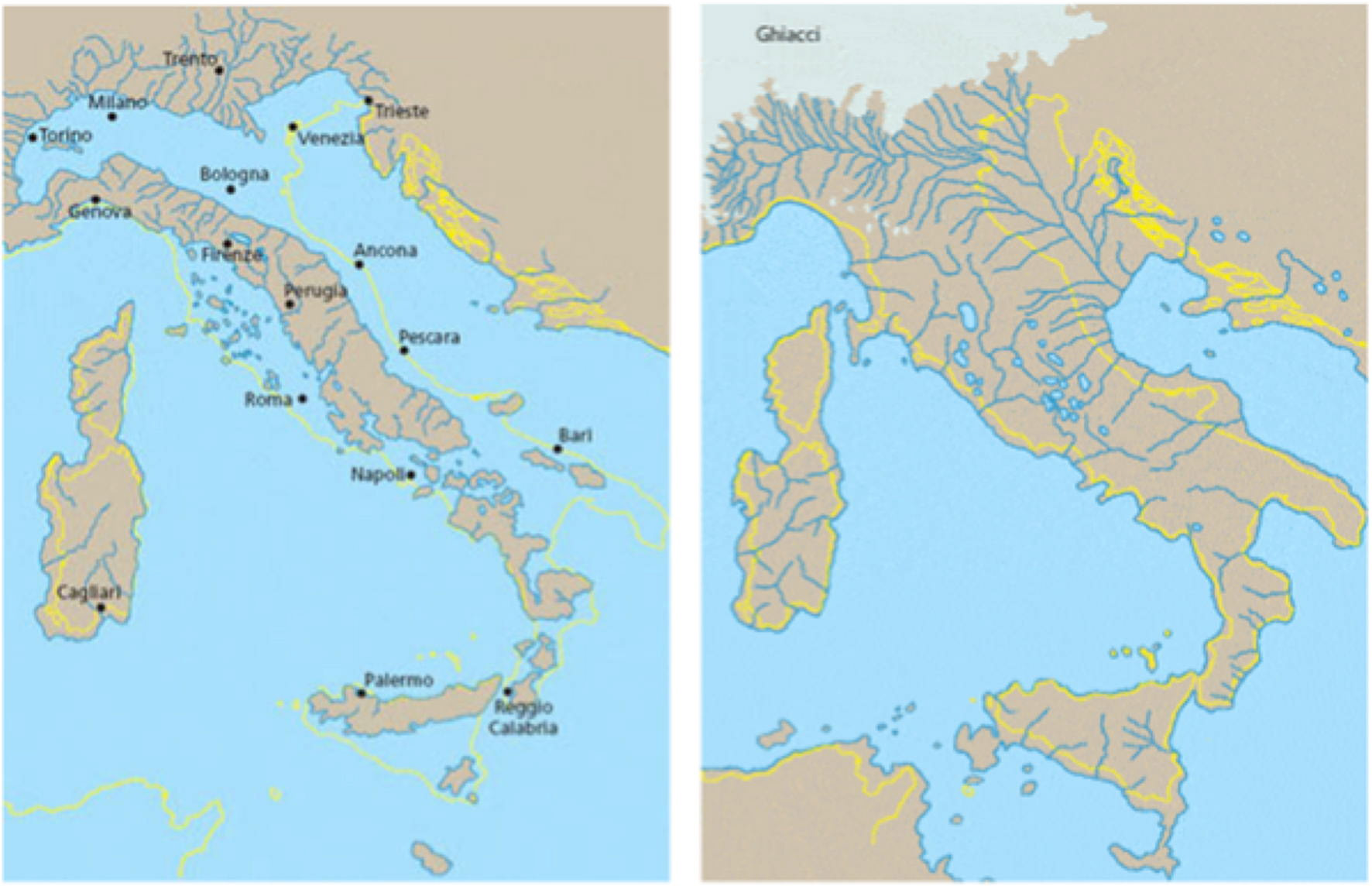
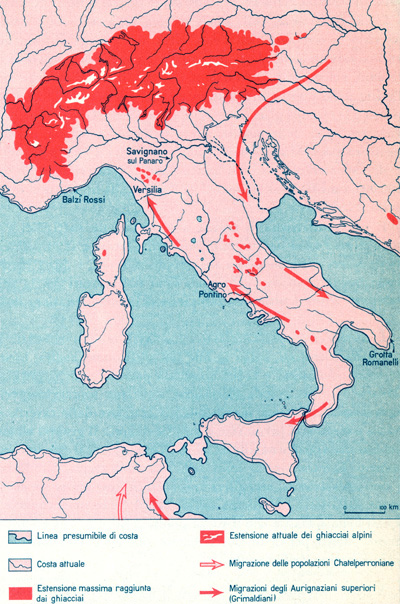 La penisola italiana durante l’ultimo periodo glaciale (Würm)
La penisola italiana durante l’ultimo periodo glaciale (Würm)
Durante l’Era Quaternaria (Pleistocene inf. e sup. – a partire da circa 3.000.000 di anni fa) le glaciazioni nell’emisfero nord divennero molto ampie con spesse coltri di ghiacci. Questo avvenne quando la placca sudamericana si saldò con quella nordamericana formando l’istmo di Panama che convoglierà la calda corrente del Golfo verso nord portando aria più umida in zone già fredde (ma prima più asciutte) e producendo le forti nevicate che si stratificheranno in spessori di migliaia di metri di ghiaccio sulle catene montuose più elevate. Premesso questo veniamo ad osservare quello che accadeva alle nostre latitudini, nella nostra penisola. Come si può osservare nell’immagine a lato l’estensione dei ghiacciai, durante l’ultima glaciazione, impediva praticamente i facili spostamenti dal centro-nord Europa verso la penisola italiana delle popolazioni di cacciatori/raccoglitori che migravano in cerca di nuovi vitali territori di caccia.
E’ da notare come a causa dell’estensione dei ghiacciai, che trattenevano una grande quantità di acqua sotto forma di ghiaccio, il livello del mar Mediterraneo era molto più basso. Infatti metà mar Adriatico era interrato, l’isola d’Elba era collegata all’attuale Toscana e la Corsica era unita alla Sardegna. La Sicilia era collegata alla Calabria e tra la Sicilia e l’attuale Tunisia, nella zona dell’isola di Pantelleria, sporgeva una lunga penisola di terra. Tutte le coste italiane erano perciò anche molto diverse e più ampie rispetto alle attuali. Le possibili migrazioni verso la penisola italiana avvennero quindi attraverso il nord Adriatico, la Puglia, la Calabria, costeggiando i bordi del mare. Alcuni importanti studiosi, suppongono che un altro possibile passaggio in migrazione possa essere avvenuto, con buona probabilità, dal nord Africa, attraverso l’attuale Tunisia, verso la Sicilia.
Tutte queste popolazioni in migrazione inizieranno a stanziarsi scontrandosi/integrandosi con i precedenti abitanti che, si pensa, già erano posizionati nella penisola italica e che probabilmente erano arrivati con una loro più antica migrazione dall’Africa orientale attraverso l’Asia Minore, il nord Africa e spingendosi prima nella penisola iberica (attraversando lo stretto di Gibilterra interrato), e poi dal sud della Francia verso l’Italia, in epoche molto più antiche.
A piena dimostrazione di tutto quanto detto, si deve citare che i più antichi reperti sia litici che ossei che sono stati ritrovati in Italia a seguito di vari scavi, in luoghi anche lontani tra di loro, e che dimostrano lo stanziamento sul nostro territorio di questi primi uomini, risalgono a:
1.000.000 Anni (Homo Erectus) Ca’ Belvedere – Monte Poggiolo(Forlì)
800.000 Anni (Homo Erectus) Cebrano – Basso Lazio
730.000 Anni (Homo Erectus) Pineta di Isernia
400.000 Anni (Homo Erectus) Quinzano – Verona
230.000 Anni (Homo Erectus-Anteneandertaliano) Balzi Rossi (Grotta del Principe) – Liguria
L’ Homo erectus, con un linguaggio fonetico molto limitato(supposizione fatta in base alla forma del cranio), viveva generalmente in piccoli gruppi a livello familiare e produceva utensili, sia litici che lignei molto semplici. Queste due condizioni gli renderanno necessario, per la sua sopravvivenza, disporre di grandi territori di caccia. La continua competizione con gli animali predatori del territorio da lui utilizzato e la perenne cacciagione necessaria alla sopravvivenza della sua piccola tribù familiare non gli permetterà di sviluppare né primitive espressioni artistiche, né forme elementari di socialità con altri gruppi paralleli.
Nella sua espansione verso nord alla ricerca di nuove zone vitali si ambienterà esclusivamente nei ripari naturali (grotte o rocce sporgenti), e circa 400.000 anni fa inizierà a dominare il fuoco. L’uso del fuoco gli migliorerà le condizioni di vita perchè potrà cuocere la carne per renderla più digeribile, scaldarsi durante la notte e difendersi meglio dagli animali predatori (orsi, tigri, leoni, iene) suoi competitori.
Per quanto riguarda la Liguria alcuni resti ossei dell’ Homo erectus sono stati rinvenuti in grotte della riviera di ponente (Balzi rossi – Comune di Grimaldi) quindi nelle vicinanze della costa marina, il che fa supporre che la sua dieta potesse comprendere anche il pesce preso direttamente sulla riva del mare, magari dopo una forte mareggiata.
Il complesso delle grotte dei Balzi rossi, che prende il nome dal colore del terreno, rossastro di origine ferrosa, si trova nella Liguria di ponente e vicino all’attuale confine francese. La non facile accessibilità all’ingresso delle grotte ha permesso la conservazione per così lungo tempo dei resti preservandoli intatti fino ai giorni nostri quando sulla fine dell’800 sono iniziate le esplorazioni in modo sistematico. Il sito è posizionato a strapiombo sul mare nel Monte Bellinda ed è formato da un nutrito complesso di antri, grotte, ripari che abitati per lunghi periodi in epoche antichissime ci forniscono così le prove certe della presenza dell’uomo preistorico.
Nella Grotta del Principe, attualmente la più grande tra le conosciute nei Balzi rossi, è stato rinvenuto il più antico resto osseo del complesso. E’ un frammento di osso iliaco destro che dopo una attenta datazione è stato attribuito ad un essere umano ANTE- NEANDERTALIANO o PRE-SAPIENS del genere HOMO ERECTUS vissuto in quella grotta 250.000 anni fa.
Per l’esame degli atri reperti di datazione più recente ritrovati in questo importantissimo complesso di grotte, si rimanda all’esame dei successivi periodi preistorici.
————————————-
GROTTA DEL PRINCIPE.
FRAMMENTO DI OSSO ILIACO DESTRO DI HOMO ERECTUS
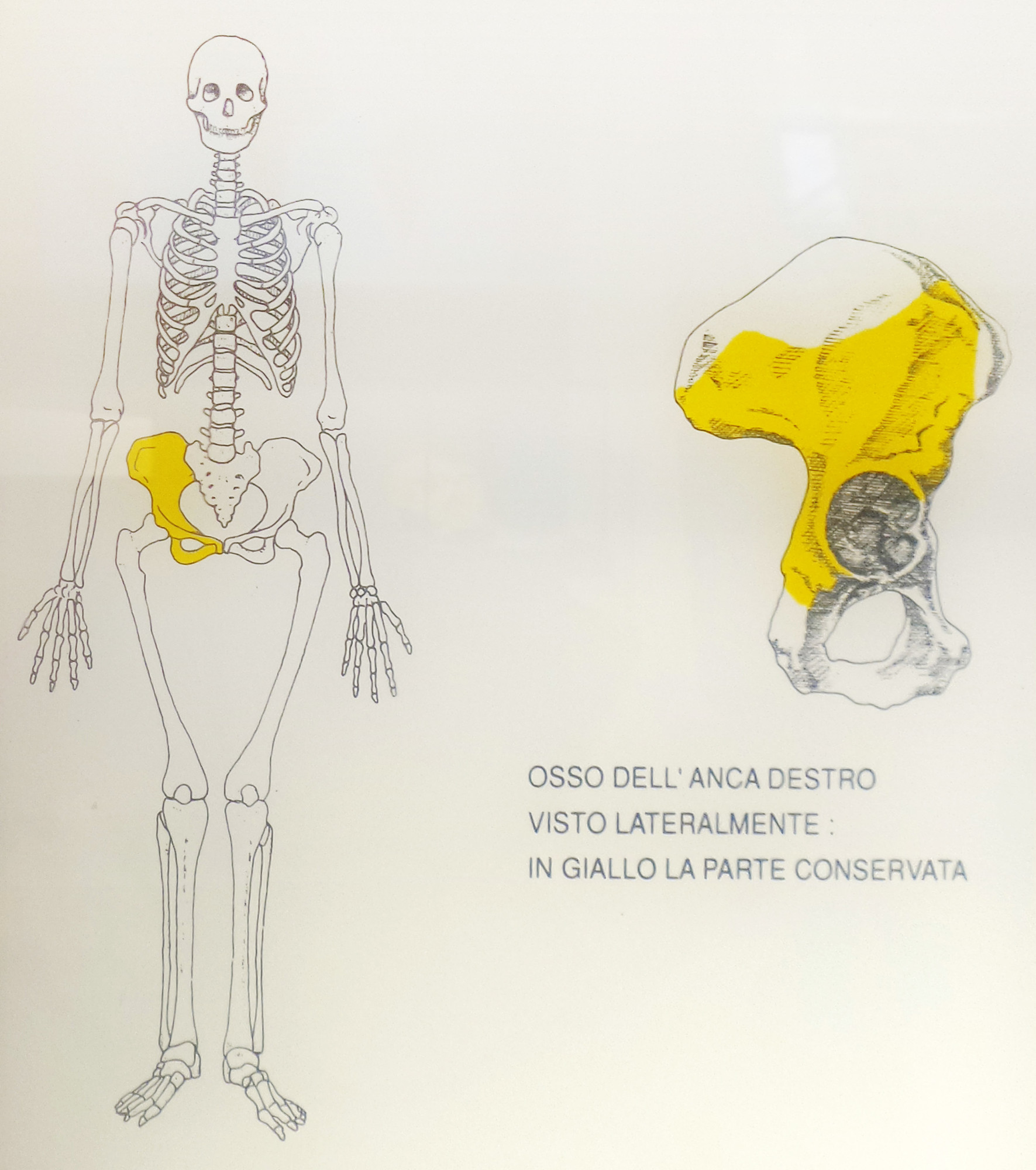
 La misura del rapporto 234U-230Th, da spettrometria gamma non distruttiva, effettuata da J.Labeyrie e Y.Yokoyama (Laboratoire des faibles radioactivités. Gif-sur-Yvette – Francia), dà un’età superiore a 230.000 anni. Di conseguenza il reperto umano, come il livello inferiore della breccia Br2, possono verosimilmente risalire al Riss II (fase fredda n°8, da 240.000 a 280.000 anni fa).
La misura del rapporto 234U-230Th, da spettrometria gamma non distruttiva, effettuata da J.Labeyrie e Y.Yokoyama (Laboratoire des faibles radioactivités. Gif-sur-Yvette – Francia), dà un’età superiore a 230.000 anni. Di conseguenza il reperto umano, come il livello inferiore della breccia Br2, possono verosimilmente risalire al Riss II (fase fredda n°8, da 240.000 a 280.000 anni fa).
Il Museo dei Balzi Rossi possiede l’osso umano più antico della Liguria e tra i più antichi d’Italia: un osso iliaco femminile di Homo Heidelbergensis, datato circa 250.000 anni fa, scoperto nella grotta del Principe dai ricercatori del Museo del Principato di Monaco in un deposito concrezionato divenuto duro come roccia. (Museo dei Balzi Rossi, www.balzirossi.it. Su concessione del Ministero dei Beni e delle Attività Culturali e del Turismo, Sopraintendenza Archeologica della Liguria.)
* * *
HOMO SAPIENS NEANDERTHAL (IN LIGURIA)
Estratto da Guido Zunino, www.vegiazena.it.
L’ HOMO di NEANDERTHAL, prende il nome da una piccola valle tra Düsserdorf ed Erberfeld nella Germania occidentale dove, per la prima volta, vennero ritrovati una calotta cranica ed alcune ossa di un ominide allora sconosciuto che aveva caratteri morfologici decisamente particolari, ma che dopo altri ritrovamenti in altre zone si dimostrerà facente parte del gruppo genetico dell’ HOMO SAPIENS.
Di corporatura non molto alta aveva le arcate sopraciliari sporgenti, calotta cranica larga e bassa, fronte sfuggente, la testa posizionata in avanti, le braccia poco più lunghe dell’attuale conformazione e le ginocchia leggermente piegate. Robusto, massiccio, muscoloso con le ossa molto spesse era geneticamente adatto all’ambiente decisamente ostile di quel periodo.
La forma del suo scheletro ricorda ancora in parte il portamento da pre-ominide delle razze primitive precedenti, anche se sono già visibili alcuni mutamenti evolutivi geneticamente più funzionali.
Il clima molto freddo ed umido e la scarsità di cacciagione lo costringevano solitario o in gruppi di due o tre uomini a lunghi percorsi di caccia su terreni impervi. Una forte muscolatura ed le ossa robuste erano quindi indispensabili alla sua sopravvivenza e conseguentemente a quella del suo nucleo familiare che dipendeva solo dalla sua predazione, se si esclude una limitata raccolta di frutti selvatici solamente durante il periodo estivo.
Il clima molto rigido di quel periodo costrinse questi uomini primitivi a scacciare gli animali che abitavano nelle grotte per insediarsi al loro posto in questi primi rudimentali ripari naturali. Seppellivano i loro morti in fosse ricoperte con terra e pietre appositamente scavate nelle stesse grotte che abitavano, spesso accanto ai loro focolari, sperando probabilmente che il calore del fuoco riscaldasse l’ormai freddo corpo del loro compagno morto.
Anche se la loro alimentazione quindi comprendeva la raccolta di frutti spontanei o di radici alimentari (come detto solo però nel periodo estivo), la loro principale fonte di vita era quindi la cacciagione che esercitavano in massa gli uomini validi del gruppo, e poiché i loro attrezzi da caccia erano molto primitivi, per riuscire a riportare indietro alla caverna una quantità di carne sufficiente alla sopravvivenza del resto del gruppo (donne, bambini, vecchi), dovevano percorrere dei lunghi tragitti, allontanandosi anche per alcuni giorni, prima del ritorno alla grotta. Questo implicava che il loro territorio di caccia-sopravvivevza era necessariamente molto ampio, e quando la selvaggina si allontanava per migrazione o per diradamento naturale, erano costretti a spostarsi in altri territori, trovandosi quindi anche una nuova caverna, cosa non sempre facile.
Vivendo ancora in piccoli raggruppamenti e dovendo anche stanziarsi su grandi spazi vitali per trovare le prede di caccia necessarie alla vita del suo gruppo, inizierà così una lenta migrazione verso nord che lo porterà dall’Africa orientale prima nel nord Africa e poi in tutta l’Europa.
Dominerà incontrastato la scena europea per almeno 250.000 anni sopravvivendo in un ambiente decisamente ostile di freddo e ghiaccio (la glaciazione di WÜRM che arriverà a ricoprire netà Asia e il centro-nord dell’Europa).
Dovrà gareggiare con animali predatori suoi rivali nella caccia, già molto scarsa alle latitudini europee durante il periodo glaciale.
Si riparerà unicamente nelle grotte e nei ripari naturali, usando inizialmente solo utensili molto semplici e rudimentali fatti di selce e legno.
Successivamente inizierà a produrre una serie di armi e di utensili: diverse varietà di asce, punte di lance e raschiatoi (INDUSTRIA MUSTERIANA dalla grotta di Le Moustier in Francia), in pratica senza variazioni per tutti i 250.000 anni di predominio sul territorio.
Conosceva ed utilizzava il fuoco sia per riscaldare le caverne, per cuocere la carne delle prede rendendola più digeribile, per allontanare gli animali feroci specialmente durante la notte o in sua assenza quando le donne e i figli restavano soli.
La sua organizzazione sociale era strutturata in piccole tribù sparse su un territorio molto ampio, e quindi molto lontane fra di loro per ragioni di spazi vitali di caccia.
Non si hanno notizie di espressioni artistiche particolari.
A conferma di quanto precedentemente detto è stato stimato che tutta la popolazione di HOMO SAPIENS – NEANDERTALIANO nel periodo di maggior diffusione (50.000 anni fa) nel sud Europa fosse allora di poco superiore ai 100.000 individui, ed intorno ai 35.000 anni fa scenderà a poche migliaia.
Solamente intorno a 35.000 anni fa si avranno delle innovazioni apprezzabili perchè dai reperti datati del periodo si iniziano a notare nuovi utensili realizzati con corna di cervi, ossa, denti di orso, e si ritrovano anche alcuni semplici monili da ornamento. Questi ritrovamenti coincidono però con il sopraggiungere in Europa dell’ HOMO SAPIENS SAPIENS “CRO-MAGNON”.
L’HOMO SAPIENS SAPIENS detto anche “CRO-MAGNON”, proveniente dall’Africa dove si era evoluto circa 200.000 anni fa, inizierà a diffondersi a partire dai 60.000 anni fa fuori dell’Africa, raggiungendo anche l’Australia riuscendo a superare alcuni tratti di mare.
In Europa giungerà nel 45.000 a.C. ed in 10.000 anni si sovrapporrà al preesistente HOMO di NEANDERTHAL, probabilmente perchè più intelligente, migliore costruttore di utensili, con una forma di linguaggio più evoluto e più versatile rispetto all’ambiente, fisicamente più alto anche se meno robusto, diventando in Europa l’HOMO di CRO-MAGNON (dalla caverna omonima in Dordogna – Francia dove sono stati rinvenuti i suoi primi resti fossili).
I più antichi resti dell’HOMO di NEANDERTHAL rinvenuti in Italia, e chiamati “L’UOMO DI SACCOPASTORE”, sono stati rinvenuti a Roma nella zona di Porta Pia e datati a 120.000 anni fa.
La presenza dell’ HOMO di NEANDERTHAL in Liguria è ampiamente documentata dai molti ritrovamenti di resti e di manufatti umani rinvenuti in varie grotte, specialmente in prossimità della costa, non perchè il territorio all’interno non fosse abitato, ma perchè probabilmente le grotte, a strapiombo sulla spiaggia e poco raggiungibili essendo a decine di metri sul livello del mare, sono rimaste intatte nei millenni successivi.
Le grotte dove sono stati ritrovate tracce o reperti attribuibili all’ HOMO di NEANDERTHAL sono diverse:
GROTTE DEI BALZI ROSSI
Il complesso delle caverne e grotte chiamate “BALZI ROSSI” (a causa del colore del terreno ricco di ossido ferroso circostante) si trova vicino al confine francese sul monte BELLINDA nel comune di GRIMALDI tra le città di VENTIMIGLIA e MENTONE a strapiombo sul mare, è composto da molti siti:
Sono state rinvenute anche diverse sepolture (doppie e singole), pitture parietali, incisioni rupestri, sculture in pietra, oggetti d’uso quotidiano ed utensili litici in pietra scheggiata di vario genere. I reperti litici ritrovati si possono attribuire al PERIODO MUSTERIANO e dimostrano quindi che le grotte erano frequentate anche nel PALEOLITICO MEDIO dall’uomo Neandertaliano.
In questi millenni, l’era glaciale, anche se in fasi altalenanti, era ancora attiva e le condizioni climatiche non facilitavano certo la vita quotidiana di questi primi uomini, ma nonostante questo, per la loro robustezza ed adattabilità all’ambiente, si diffusero e si stanziarono sul territorio, precedendo la successiva migrazione dell’ HOMO SAPIENS SAPIENS chiamato anche di CRO-MAGNON che si sovrapporrà sugli stessi territori.
GROTTE DI FINALE LIGURE
Le grotte che si trovano nel territorio di FINALE LIGURE in provincia di SAVONA sono state abitate già in epoca molto antica ed attribuibile al periodo che stiamo esaminando, e sono principalmente:
La caverna delle ARENE CANDIDE, che prende il nome da una duna di sabbia silicea chiara (trovata all’epoca dell’esplorazione) presso il promontorio della CAPRAZOPPA, si trova a circa 90 metri sul livello del mare presso Finale Ligure.
Nella caverna delle FATE, anch’essa presso Finale Ligure, sono stati rinvenuti esclusivamente Manufatti Litici attribuibili all’UOMO di NEANDERTHAL del PALEOLITICO MEDIO (databili tra 100.000 e 50.0000 anni fa) più precisamente attribuibili ai periodi chiamati LEVALLOSIO e MUSTERIANO.
Ma non essendo stati trovati resti umani o necropoli di alcun genere è possibile che o non fosse ancora in uso in quella zona la sepoltura o che venisse utilizzata per l’uso un’altra caverna (come abbiamo già visto sarà fatto in epoca successiva nella caverne delle Arene Candide).
Le GROTTE di TOIRANO si trovano nel primo entroterra tra Albenga e Savona nel ponente ligure e sono un complesso di grotte che si inoltrano nel cuore della montagna con percorsi ed andamento molto vario, formate o da stretti cunicoli o da grandi saloni, molto ricche anche di stalattiti e stalagmiti.
Con certezza esse sono state frequentate sia dall’uomo primitivo che da molti animali selvatici come l’orso delle caverne, come risulta dai ritrovamenti dei resti di entrambi.
Probabilmente sia l’orso che l’uomo erano a caccia o di un riparo sicuro o di prede animali ed hanno frequentato entrambi la stessa grotta. Non è accertato se questo avveniva in periodi diversi o contemporaneamente.
Nella GROTTA delle STREGHE infatti, vicino a resti ossei dell’Orso delle Caverne, si può chiaramente vedere impressa nel fango allora molle ed ora pietrificato, una impronta di un piede perfettamente conservata di un giovane Neandertaliano ( HOMO SAPIENS – NEANDERTHALENSIS databile a circa 30.000 anni fa).
L’ HOMO di NEANDERTHAL si estinguerà quindi naturalmente tra i 28.000 e i 30.000 anni fa perchè probabilmente non in grado di competere con l’intelligenza organizzativa, i gruppi tribù più numerosi e l’abilità manuale e di caccia dell’ HOMO SAPIENS SAPIENS (CRO-MAGNON) che darà origine quindi alle genti che abiteranno l’Europa e l’Asia in seguito e da cui appunto si evolveranno i prototipi delle varie razze attualmente viventi.
Gli ultimi resti, in ordine di datazione, attribuibili con certezza all’ HOMO di NEANDERTHAL sono stati rinvenuti nella Spagna meridionale, nell’Italia centrale, nella Croazia.
Da una approfondita indagine eseguita sul DNA dei reperti fossili delle due tipologie risulta che le due razze non sono compatibili quindi si può supporre che non si fusero, anche se coabitarono per molto tempo.
L’unica notizia che dovrebbe documentare l’avvenuta fusione tra le due genie dipenderebbe dal ritrovamento che nel 1998 è stato fatto nella valle del LEPIDO in Portogallo, dove si è rinvenuto uno scheletro di un bimbo di 4 anni che riporta le caratteristiche miste delle due specie, e che alla datazione risulterebbe essere vissuto in un’epoca stimata a circa 3.000 anni dopo la presunta estinzione dell’ HOMO di NEANDERTHAL.
* * *
L’HOMO SAPIENS NEANDERTHAL AI BALZI ROSSI
Cartelli e parte delle immagini sono state fatte al Museo dei Balzi Rossi. Le riprese fotografiche sono state effettuate su concessione del Ministero dei Beni e delle Attività Culturali e del Turismo, Sopraintendenza Archeologica della Liguria.
Cartello 3
Benché situati in una zona isolata, i giacimenti preistorici dei Balzi Rossi non sono giunti intatti ai primi studiosi che vi eseguirono scavi.
Già nel 1786 il naturalista H.B. de Saussure, che segnalò pure la presenza di fori di litodomi sulla parete rocciosa, notava l’esistenza di un forno da calce nella Grotta dei Fanciulli, mentre un altro è ricordato successivamente dal Forel e dal Rivière nella Grotta del Caviglione.
La messa in opera di queste strutture deve certamente aver portato alla luce considerevoli quantità di industria litica e ossa di animali.
Pur essendo dunque le grotte sicuramente note da tempo agli abitanti della zona per questo aspetto, il primo a mostrare un interesse di tipo scientifico sembra sia stato il principe Florestano I di Monaco, che a partire dal 1846 fece raccogliere una campionatura di materiali, in particolare dalla caverna che porta il suo nome, ritenendoli degni di studio.
Tra il 1854 e il 1858 A.Grand di Lione eseguì alcune campagne di scavo nelle grotte, raccogliendo industria litica e constatando che gli oggetti più significativi si incontravano ad una certa profondità al disotto del deposito argilloso che ne formava il suolo.
I primi scavi che dedicarono un certa attenzione alla descrizione dei depositi furono però quelli compiuti nel 1858 da F.Forel, presidente della Società Storica della Svizzera Romanda, che concentrò il proprio interesse sulla Grotta del Caviglione e sulla Barma Grande. Forel annotava la distanza dei suoi ritrovamenti rispetto alla volta della grotta e la profondità delle sue trincee. Nella Grotta del Caviglione incontrò, a dieci metri dalla volta, uno strato, ricco di resti ossei di animali, cenere e carboni di un metro e mezzo di potenza, sovrastante un livello sterile. Nella Barma Grande, in cui allora il livello del suolo era a quasi quindici metri dalla volta, incontrò invece uno strato superficiale con industria litica di circa 50 centimetri, al disotto del quale vi era nuovamente uno strato sterile. Tra i reperti faunistici ritenne di individuare la presenza di numerose specie, tra cui Cervo, Capriolo, Stambecco, Uro, Cavallo, Cinghiale, Lupo, Volpe, Gatto selvatico, Coniglio e un Cetaceo.
Questi primissimi scavi segnalarono al mondo scientifico, che dibatteva allora il problema dell’antichità dell’Uomo, l’esistenza di depositi preistorici ai Balzi Rossi.
Nello stesso anno altre ricerche furono compiute dal prof. A.Perez e da P.Geny per incarico del governo piemontese. Tra i materiali raccolti e consegnati al Museo di Stona Naturale di Genova, oltre ad utensili paleolitici figurano anche tre asce neolitiche in pietra levigata, che diedero luogo a non poche discussioni, ma che provengono invece da Nizza, come fu successivamente possibile accertare.
Tra il 1862 ed il 1871 fu attivo ai Balzi Rossi P.G.S.Moggridge, che scoprì un focolare con abbondante industria litica nel Riparo Lorenzi e scavò nella Grotta del Caviglione per 60 centimetri al disotto del focolare più alto, mettendo in luce cinque strati con resti di frequentazione antropica.
Altri sondaggi furono condotti nel 1864 da E.Chantre, sempre nella Grotta del Caviglione.
Negli stessi anni e fino al 1871 S.Bonfils, un appassionato cultore di scienze naturali di Mentone, visitava assiduamente le grotte, raccogliendo un’interessante collezione di resti scheletrici di animali, conchiglie ed industria litica e su osso, che formarono il primo nucleo di materiali del Museo Municipale di Mentone da lui fondato. In particolare condusse diverse campagne di scavo nella Barma Grande, segnando preventivamente il limite superiore del deposito con una fila di chiodi, la cui traccia si può tuttora scorgere.
In esito a tale attività pubblicò, insieme a L.Smyers, un volume in cui esaminava l’industria litica raccolta tentando di comprendere come fossero fabbricati e per quale uso i diversi utensili: Recherches sur les outils eri silex des Troglodytes et sur la manière dont ils les fabriquaient, Nice 1872.
Minori interventi furono compiuti dal prof.Broca nel 1865 e dal conte Costa de Beauregard nel 1868, ancora nella Grotta del Caviglione.
Queste indagini, benché abbiano avuto il merito di far conoscere le grotte dei Balzi Rossi come giacimenti preistorici, causarono con la loro carenza di metodo e la loro disorganicità, dovuta anche al succedersi di molti scavatori, la distruzione di parti importanti dei depositi, di cui è rimasta una documentazione minima. I materiali raccolti furono inoltre dispersi in numerose collezioni, sia private che di istituti culturali, tanto che se ne conoscono lotti destinati a Siena, Torino, Lione, Nizza, Ginevra, Genova e Mentone.
——————————
[in costruzione]
GROTTE DI FINALE LIGURE
GROTTA DELLE ARENE CANDIDE
——————————
[in costruzione]
GROTTE di TOIRANO
GROTTA DELLE STREGHE
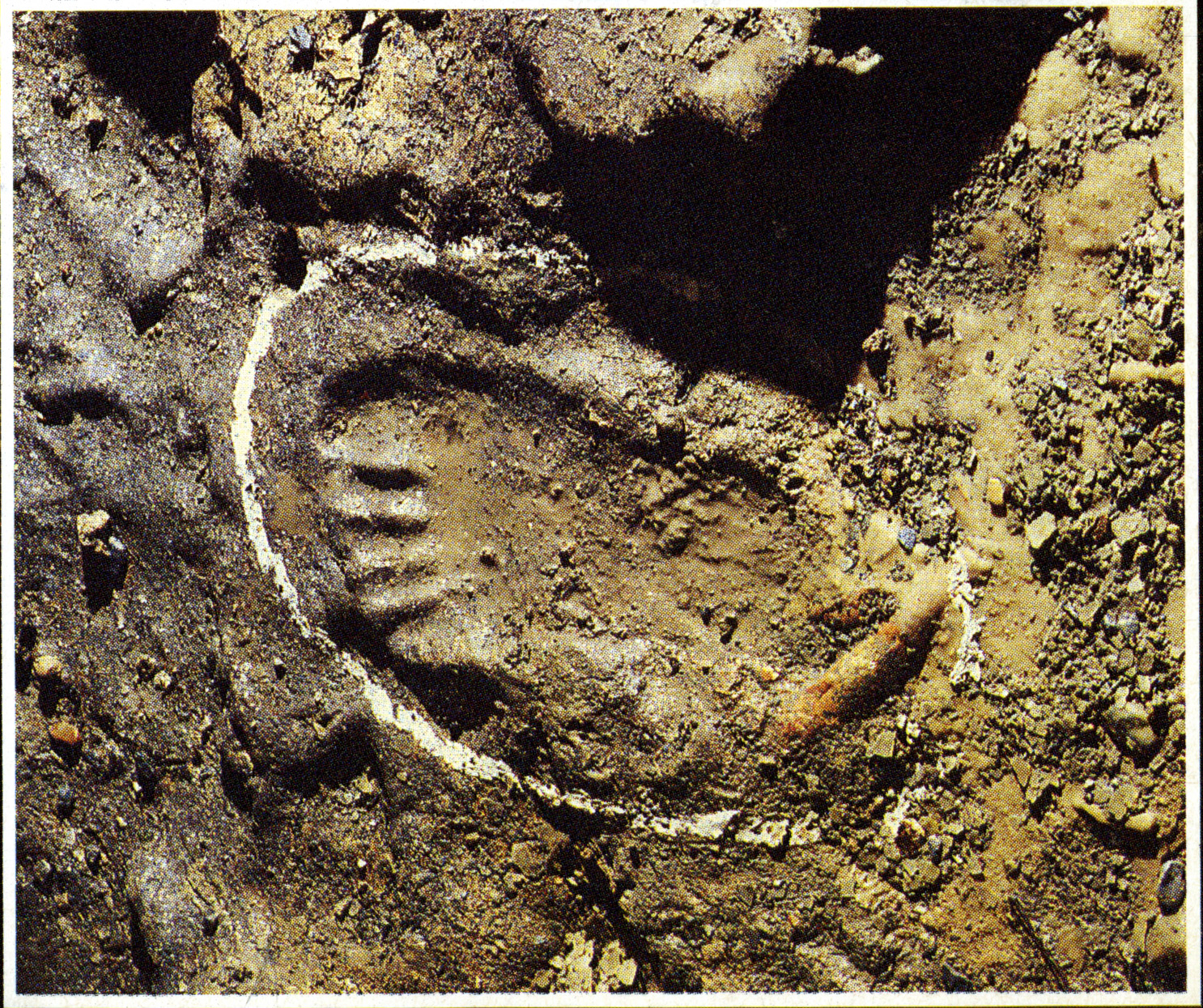
[inserire testo Toirano]
* * *
FAUNA NEL PALEOLITICO
Cartelli e alcune immagini sono state fatte al Museo dei Balzi Rossi. Le riprese fotografiche sono state effettuate su concessione del Ministero dei Beni e delle Attività Culturali e del Turismo, Sopraintendenza Archeologica della Liguria.
Cartello 8
LA FAUNA
LA MALACOFAUNA DELLA SPIAGGIA TIRRENIANA
Il Quaternario è caratterizzato dall’alternarsi di periodi freddi (glaciali) e periodi più caldi (interglaciali). Queste fasi climatiche provocavano variazioni del livello marino, con conseguente formazione di spiagge a livelli diversi da quello attuale. Nei periodi glaciali si ha nel Mediterraneo un abbassamento del livello marino che causa la formazione di spiagge attualmente sommerse, nei periodi interglaciali invece il livello marino si alza e si formano linee di spiaggia al di sopra di quella attuale.
Le spiagge riferite al Tirreniano corrispondono ad una fase calda che si articola in due piani, riferibili all’interglaciale Mindel-Riss (Tirreniano I) e all’interglaciale Riss-Würm (Tirreniano II), separati da una fase regressiva corrispondente alla glaciazione rissiana. Il Tirreniano II si conclude con una forte regressione marina corrispondente all’inizio della glaciazione wurmiana
Durante questo periodo compaiono nel Mediterraneo alcuni molluschi di ambiente caldo, viventi attualmente sulle coste del Senegal, tra cui Strombus bubonius, Conus guinalcus, Natica lactea, Mytilus senegalensis e Cardita senegalensis, accanto ai quali vivevano numerose specie che troviamo anche attualmente nel Mediterraneo.
Il raffreddamento conseguente all’inizio della glaciazione wurmiana provocò la scomparsa di questi molluschi, mentre continuarono a sopravvivere quelli più adatti ad un ambiente temperato, molti dei quali presenti nel Mediterraneo sin dalla fine del Terziario, superando le precedenti fasi fredde.
Le conchiglie provenienti dalla spiaggia tirreniana dei Balzi Rossi appartengono in gran parte a specie di scogliera (Astraea rugosa, Conus mediterraneus, Patella ferruginea, Spondylus gaederopus), ma sono presenti anche specie che vivono nella sabbia (Glycymeris violacescens, Callista chione, Rudicardium tuberculatum). Doveva quindi trattarsi di un ambiente che alternava zone rocciose a tratti sabbiosi. Lo Strombus bubonius è abbastanza frequente.
LA FAUNA TERRESTRE DEL PALEOLITICO
Anche sulla terraferma durante il Pleistocene, a causa dell’alternarsi di fasi glaciali e interglaciali, si ha un susseguirsi di ambienti ed associazioni faunistiche che si adattano alle condizioni climatiche del momento. Inoltre nei periodi interglaciali, con l’innalzamento del livello marino, vengono sommersi tratti di costa, che invece restano liberi nei periodi più freddi, quando il mare è in fase regressiva.
In Italia si hanno ritrovamenti di ossa di animali non derivanti da attività umana per il Quaternario inferiore (giacimenti del Valdarno, di Olivola ecc.) e altri giacimenti più recenti che hanno restituito una grande quantità di resti animali associati ad industria di tipo Paleolitico inferiore ad Isernia La Pineta (circa 730.000 anni da fa) e Venosa. Resti attribuiti alla glaciazione rissiana sono stati trovati anche nella Grotta del Principe e in altre località come Castel di Guido (Roma), Torre in Pietra e Loreto di Venosa.
Nel Paleolitico inferiore troviamo tra gli animali di ambiente terrestre alcune specie di grandi dimensioni, come l’Elefante antico, oltre a rinoceronti, ippopotami e bisonti. Si affermano anche altri animali come l’Uro, il Cavallo e alcuni Cervidi che continueranno ad esistere per un lungo periodo. Tra i carnivori sono stati trovati alcuni Canidi, Ienidi, Ursidi e grossi Felini.
Ai Balzi Rossi, dopo la fase di trasgressione marina che causò il formarsi della spiaggia tirreniana, le grotte e in seguito la fascia costiera furono liberate dal mare e immediatamente occupate dall’Uomo e dai grossi mammiferi terrestri. I resti di pasto che troviamo subito al di sopra della spiaggia sono ancora costituiti da ossa di animali di clima caldo, come il Rinoceronte di Merck, l’Elefante antico, l’Ippopotamo, la Iena, il Leopardo. Oltre a questi animali tipicamente “caldi” vivevano però già da tempo in Italia, come si è visto, altre specie più adattabili, che si ritrovano poi anche in periodi con clima freddo o temperato, come l’Uro, il Cavallo, il Cervo ecc.
Con il progredire della glaciazione würmiana, gli animali di ambiente caldo diventano più rari e vengono in seguito sostituiti da specie in grado di sopportare un clima più rigido. Nella fase più antica del Würm si trovano ancora resti di Rinoceronte di Merck, Leopardo, Leone, Elefante antico, Ippopotamo e Iena accanto ad animali di clima freddo come lo Stambecco, il Camoscio e la Marmotta. Nel Würm II l’Elefante antico e l’Ippopotamo tendono a scomparire, mentre comincia a trovarsi il Mammut, di clima traodo, e diventa molto comune l’Orso speleo. Ai Balzi Rossi troviamo in questo periodo una maggioranza di animali di foresta, come il Cinghiale e il Cervo, accanto a resti di Cavallo che testimoniano l’esistenza di una zona di steppa. Il raffreddamento del clima è indicato dalla presenza dello Stambecco.
La differenziazione tra le associazioni faunistiche è causata, oltre che dal clima, dalle caratteristiche ambientali. Così l’Elefante, il Rinoceronte, il Cavallo sono caratteristici di un ambiente aperto, di steppa, mentre, nel caso di grotte situate in ambiente montano e forestale si potrà avere un numero superiore di resti di Cervidi, oltre a carnivori come l’Orso e il Leone.
Ai Balzi Rossi l’abbondanza di animali di steppa poteva essere favorita dalla presenza di una pianura davanti alle grotte. L’attività dell’Uomo, che poteva cacciare in diverse aree, può provocare l’associazione, nello strato archeologico, di resti di animali con esigenze diverse.
Nel Paleolitico superiore scompare ogni traccia degli animali di clima caldo, mentre le associazioni faunistiche si evolvono in senso attuale. Il clima freddo favorisce il proliferare anche a bassa quota di specie come il Camoscio, lo Stambecco, la Marmotta mentre,continuano e si affermano il Cinghiale, il Cervo, il Capriolo, il Daino, l’Uro, il Cavallo e l’Asino idruntino. Ai Balzi Rossi è stata trovata anche la Renna, che manca nelle altre regioni italiane, mentre è comune oltre le Alpi. Tra i carnivori troviamo specie tipiche di foresta come la Lince, il Gatto selvatico, la Martora, ed altre di ambiente più vario come il Lupo e la Volpe. L’Orso bruno tende, nei periodi più recenti, a sostituire l’Orso speleo.
La selezione operata dall’Uomo nella formazione dei depositi è in genere abbastanza forte e alcune specie possono essere rappresentate da un gran numero di ossa perchè più facilmente cacciabili o più richieste per particolari usi come la lavorazione dell’osso o del corno o per le pelli. Oltre che dai resti delle ossa si conoscono gli animali di questo periodo anche dalle raffigurazioni dipinte o scolpite che sono state trovate in alcune grotte, soprattutto in Francia e Spagna, ma attestate anche ai Balzi Rossi (equide della Grotta del Caviglione). Tra gli animali più raffigurati si possono ricordare l’Uro, il Bisonte, lo Stambecco, il Cavallo, il Mammut, il Rinoceronte, ma non mancano animali di piccola taglia come pesci, uccelli o piccoli mammiferi.
Il motivo di queste raffigurazioni è incerto, in genere viene loro attribuito un significato magico, giustificato in alcuni casi dalla presenza di armi o di mani in prossimità degli animali.
Con la fine della glaciazione würmiana gli animali più tipicamente “freddi” come lo Stambecco, il Camoscio e la Marmotta vengono respinti quasi ovunque dalle specie che si adattano al clima temperato come il Cervo, il Capriolo, l’Uro, il Cinghiale e l’Orso bruno, e il loro habitat si restringe alle zone alte delle montagne. Altri animali come il Cavallo, l’Asino idruntino e il Daino tendono a scomparire dalle nostre regioni nel postglaciale, resistendo più o meno a lungo in alcune aree più favorevoli.
 Fig. 1. Malacofauna dalla spiaggia tirreniana situata alla base del deposito della Grotta del Principe (scavi de Villeneuve): Strombus bubonius (a sinistra) e Patella ferruginea.
Fig. 1. Malacofauna dalla spiaggia tirreniana situata alla base del deposito della Grotta del Principe (scavi de Villeneuve): Strombus bubonius (a sinistra) e Patella ferruginea.
Fig. 2. Mandibola di Rinoceronte di Merck dalla Grotta del Principe (scavi de Villeneuve).
Fig. 3. Presenza delle principali specie di mammiferi ai Balzi Rossi durante il Paleolitico inferiore, medio e superiore. La linea nera alla sommità delle colonne indica l’estinzione della specie o la sua scomparsa dalla zona alla fine del Pleistocene.
Cartello 9
L’AVIFAUNA DELLA BARMA GRANDE
Un campione di resti di avifauna è stato raccolto nel 1949 alla Barma Grande, nell’area in cui si trovavano le vetrine distrutte nel corso della Seconda Guerra Mondiale.
Le differenze di abitudini e di ambiente delle specie identificate sono probabilmente da collegare alle differenze climatiche che caratterizzavano le varie fasi del Paleolitico in cui si è formato il deposito della grotta.
Sono numerose le specie legate alle acque dolci, molte delle quali tuttavia possono essere trovate anche in mare, soprattutto durante le migrazioni (Svasso maggiore, Oca lombardella, Fischione, Alzavola, Germano reale, Marzaiola, Orchetto marino) mentre altre sono più strettamente legate alle acque interne (Moriglione, Moretta, Smergo maggiore, Tarabuso, Folaga, Chiurlo, Gufo di palude). Tipici uccelli marini sono il Marangone dal ciuffo e il Mugnaiaccio, mentre altri nidificano di preferenza su scogliere e dirupi, anche vicino al mare (Pellegrino, Aquila reale, Piccione selvatico, Rondone, Rondine, Taccola e Corvo imperiale); spesso si trova sulle rive del mare anche l’Albanella reale. Sono indicatori di ambiente diverso dall’attuale il Fagiano di monte e il Gracchio, che attualmente vivono ad alta quota, e un gruppo di specie di steppa e pietraia (Coturnice, Pernice rossa, Starna, Quaglia, Re di quaglie. Gallina prataiola).
Altre specie sono legate al bosco (Biancone, Sparviero, Barbagianni, Civetta, Allocco, Ghiandaia), mentre il Lanario nidifica sulle scogliere in genere lontano dal mare e le altre specie possono adattarsi a diversi ambienti (Poiana, Merlo, Gazza, Cornacchia).
LE VERTEBRE DI SALMONIDI DELLA TRIPLICE SEPOLTURA
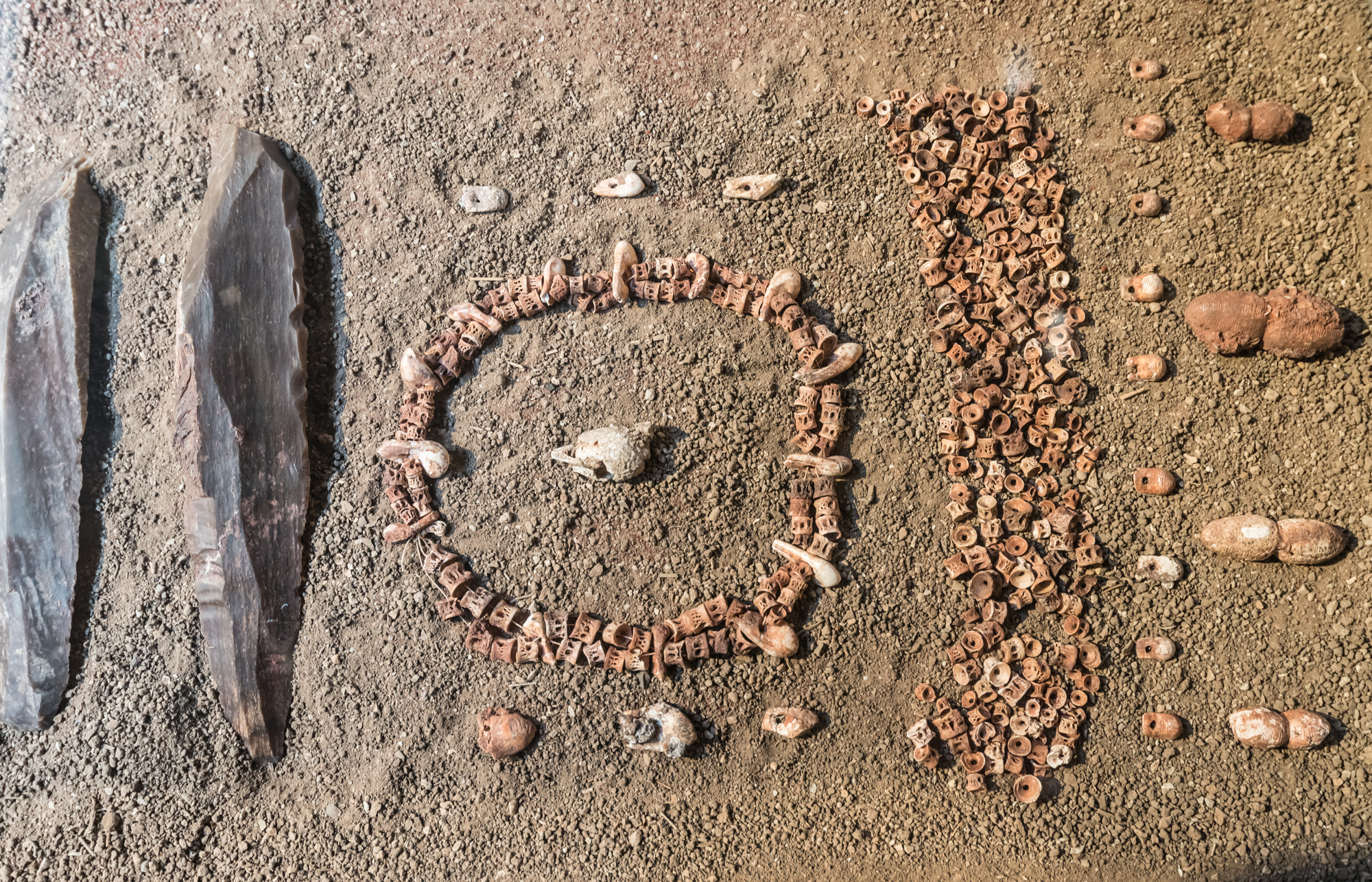 Rinvenute al momento dello scavo sul cranio di tutti e tre gli individui e presso il collo di uno dei giovani, le vertebre studiate sono in totale 244 e, premesso che la distinzione tra Salmo trutta (Trota) e Salmo salar (Salmone) risulta difficoltosa per le somiglianza tra le due specie, si può concludere che si tratti con ogni probabilità di Trota (Salmo trutta L.).
Rinvenute al momento dello scavo sul cranio di tutti e tre gli individui e presso il collo di uno dei giovani, le vertebre studiate sono in totale 244 e, premesso che la distinzione tra Salmo trutta (Trota) e Salmo salar (Salmone) risulta difficoltosa per le somiglianza tra le due specie, si può concludere che si tratti con ogni probabilità di Trota (Salmo trutta L.).
Per quanto riguarda i caratteri esterni, le vertebre toraciche della Trota hanno, come quelle esaminate, profilo più concavo di quelle del Salmone dal punto di vista dorsale; tale somiglianza è confermata anche dall’esame radiografico. Dall’esame degli anelli di accrescimento risultano 16 vertebre di pesci morti in inverno e una di individuo morto in primavera: si può quindi supporre che le trote venissero pescate durante la risalita dei fiumi. Si tratterebbe in questo caso di Trota marina, come probabile anche per la grossa taglia delle vertebre. La Trota marina raggiunge una lunghezza di circa un metro (quella di fiume non supera i 50 cm) e vive in acqua dolce fino ad un massimo di cinque anni; dopo uno o due anni di vita in mare risale i fiumi per riprodursi, anche più volte a differenza del Salmone.
La Trota non vive attualmente nel Mediterraneo a causa della salinità troppo elevata, ma la sua presenza è segnalata nel Paleolitico superiore alla Grotta della Madonna di Praia a Mare.
Secondo J.J.Cleyet-Merle i Salmonidi costituivano da soli il 37,8% dei pesci identificati dal Maddaleniano all’Epipaleolitico; questa abbondanza può essere spiegata proprio con la facilità della pesca durante la risalita dei fiumi o durante la successiva discesa, quando molti individui si possono raccogliere anche morti.
Nel campione esaminato sono presenti esclusivamente vertebre toraciche, probabilmente a causa dell’ assenza di apofisi saldate, che le rendevano già pronte per essere utilizzate.
Da un esame approfondito risulta che venivano forate per pressione ripetuta di una punta sottile. La maggior parte presenta la superficie esterna resa lucida dall’uso, ma raramente la zona di contatto tra due vertebre è usurata, cosa che fa supporre che non fossero molto ravvicinate fra loro.
Sembra però che almeno una parte non fosse cucita singolarmente su un’eventuale base di altro materiale, essendo alcune unite da concrezioni, come se fossero state legate da un filo una di seguito all’altra.
LA LAVORAZIONE DELL’OSSO, DELL’AVORIO E DEL CORNO
A partire dal Paleolitico superiore si hanno numerose testimomanze sulla lavorazione dell’osso, ma questo materiale era usato per la fabbricazione di strumenti già nel Paleolitico inferiore (Castel di Guido presso Roma).
Le ossa venivano ricavate dagli ammali normalmente consumati per l’alimentazione. Si nota una scelta di determinate ossa per i diversi tipi di strumenti. Tra quelle più usate si trovano i metapodi, e in genere le ossa lunghe degli arti, e le coste. La lavorazione dell’osso continua oltre il Paleolitico anche in periodi più recenti, e la sua importanza diminuisce solo in periodo storico. I denti venivano spesso utilizzati anche come oggetti ornamentali: sono frequenti, dal Paleolitico superiore, i canini superiori di cervo forati.
L avorio, che poteva essere ottenuto dai denti di alcuni grossi animali (Elefante antico, Mammut, Ippopotamo), veniva in genere adoperato per la fabbricazione di oggetti ornamentali o cultuali (pendagli, statuette), più raramente per utensili. L’uso dell’avorio decade con la scomparsa dall’ Europa degli animali da cui si ricavava e solo molto più recentemente, in età protostorica, con lo sviluppo dei commerci ricompaiono in Italia oggetti di avorio.
Oltre a questi materiali venivano largamente impiegate, dal Paleolitico superiore, le corna dei Cervidi. Le corna che in questi ammali cadono e vengono sostituite ogni anno, venivano raccolte anche dopo la caduta. Dalle corna si ricavavano utensili, armi ed oggetti ornamentali Nel caso di corna ricavate da animali uccisi, si usava in genere staccarle tagliandole al di sotto della rosetta. Ciascun corno veniva poi suddiviso in porzioni lavorabili.
Un materiale del tutto diverso si ricavava invece dalle corna dei Bovidi. In questi animali le corna sono formate da una struttura ossea portante (cavicchia) e da un astuccio corneo che non si conserva nei depositi archeologici. Un indizio che anche questo materiale venisse usato dall’Uomo è dato dai segni di tagli provocati, durante il distacco dell’astuccio corneo, sulla cavicchia ossea. L’ astuccio poteva essere usato intero come recipiente o tagliato per fabbricare oggetti vari. L’ uso di questo materiale, anche se poco documentato, era molto esteso durante la preistoria e anche in epoca storica.
Oltre ai segni lasciati sulle ossa dalla lavorazione intenzionale da parte dell’Uomo, altri segni possono essere dovuti alle pratiche di macellazione e alle tracce di denti di animali.
I tagli e i graffi di macellazione sono più o meno profondi e il loro studio permette di ricostruire le tecniche usate per scuoiare e tagliare a pezzi le prede. Sono dovuti alla scuoiatura i graffi sul cranio, sulla mandibola e sulle parti estreme degli arti. Le ossa venivano poi disarticolate per tagliare le porzioni e spezzate per recuperare il midollo. Il distacco della carne dalle ossa produceva una serie di graffi sulla diafisi. In alcuni periodi le grotte furono abbandonate dall’Uomo e occupate dagli animali, sopratutto Iene che vi lasciarono i resti delle loro prede.
I segni lasciati sulle ossa da animali si possono attribuire a due gruppi principali: quelli lasciati dai carnivori, consistenti in una serie di impressioni tonde, e quelli lasciati dai roditori, caratterizzati da una serie di solchi paralleli.
 Fig. 1. La fauna della zona dei Balzi Rossi prima della glaciazione del Würm, durante una fase temperato-calda. Si noti la presenza di numerosi pachidermi e carnivori.
Fig. 1. La fauna della zona dei Balzi Rossi prima della glaciazione del Würm, durante una fase temperato-calda. Si noti la presenza di numerosi pachidermi e carnivori.

Fig. 2. La fauna della zona dei Balzi Rossi all’inizio della glaciazione del Wurm. Il clima abbastanza temperato permette ancora la presenza dell’Elefante Antico e del Rinoceronte di Merck.

Fig. 3. La fauna della zona dei Balzi Rossi durante le fasi più fredde della glaciazione del Würm. Si noti la presenza di animali di clima freddo come lo Stambecco, la Renna, il Camoscio e la Marmotta.
* * *